

[et_pb_section fb_built=”1″ _builder_version=”3.22″ custom_padding=”0px|0px|0px|0px”][et_pb_row make_equal=”on” _builder_version=”3.25″ background_size=”initial” background_position=”top_left” background_repeat=”repeat” custom_padding=”27px|0px|0|0px” z_index_tablet=”500″][et_pb_column type=”4_4″ _builder_version=”3.25″ custom_padding=”|||” custom_padding__hover=”|||”][et_pb_text admin_label=”Resumen” _builder_version=”3.29.3″ header_text_color=”#35568a” header_2_text_align=”left” header_3_text_color=”#35568a” header_5_text_color=”#1a9bc2″ custom_margin=”||0px|” z_index_tablet=”500″]
Los micronutrientes son compuestos orgánicos necesarios en pequeñas cantidades para los vertebrados, aunque muy esenciales para mantener las funciones metabólicas (Martins et al., 2016). Dentro de los micronutrientes se encuentran las vitaminas y los minerales. La mayor parte de las vitaminas no son sintetizadas en las células animales y en los rumiantes deben ser abastecidas por las dietas. En algunos casos, su síntesis en el rumen puede no ser suficiente para el máximo crecimiento y desempeño de la producción de estos animales (Wu, 2018).
Se ha demostrado que la mayor parte de nuestros pastos, especialmente en la sierra del país, son deficientes en nutrientes. En Australia, una zona con abundantes pastos aparentemente nutritivos, Knowles y Grace (2016) encontraron que las pasturas no proporcionaban suficientes nutrientes para satisfacer los requerimientos dietéticos de los animales, con el 44% de los pastos siendo nutritivamente deficientes. En Perú, la mayor parte de los rumiantes son criados al pastoreo y deben obtener sus nutrientes de los forrajes disponibles, que varían ampliamente por zona, altitud y época del año.
Por otro lado, aunque la dieta en base a forraje verde puede proporcionar suficiente cantidad de nutrientes, la suplementación intensiva con alimento almacenado en forma de heno o ensilado puede reducir hasta en 50% los niveles de nutrientes en los alimentos (Martins et al., 2016). También, los rumiantes alimentados con una dieta que contiene poco o ningún forraje verde están en riesgo de desarrollar signos clínicos de deficiencia de vitaminas y minerales. Se ha reportado que también ocurre deficiencia en animales alimentados con dietas cuyos suplementos minerales y vitamínicos están vencidos (Kang et al., 2017).
Los principales grupos en riesgo de sufrir de deficiencia de nutrientes son las hembras en periparto y los neonatos, en especial cuando nacen en períodos de cambios climáticos o cuando la oferta de forraje verde disminuye. También están en riesgo los animales criados en estabulación, cuando son alimentados principalmente con concentrados que no tienen la suplementación correcta de vitaminas y minerales (Martins et al., 2016), o cuando los sacos de concentrado son expuestos a luz intensa y calor, lo cual acelera la destrucción de algunos nutrientes en los suplementos (Kang et al., 2017).
Funciones de algunos micronutrientes esenciales
Calcio. El calcio es el mineral más abundante en el organismo animal. Es un constituyente importante del esqueleto y los dientes, en los cuales es hallado casi el 99% del calcio corporal. Además, es un constituyente esencial de las células y fluidos tisulares. El calcio es esencial para la actividad de una cantidad de sistemas enzimáticos, incluyendo aquellos necesarios para la transmisión de los impulsos nerviosos, para la contractibilidad del músculo y para la producción láctea. También participa en la coagulación sanguínea ya que es un elemento presente en el plasma, alrededor de 80-120 mg por litro (McDonald et al., 2010). Las principales fuentes de calcio para los animales son los cereales y las raíces (Wu, 2018).
Vitamina A. La vitamina A tiene múltiples funciones. Excepto para la visión, donde se utiliza retinol, la mayoría de acciones fisiológicas de la vitamina A son mediadas por su metabolito bioactivo, el ácido retinoico. El ácido retinoico participa en la síntesis de glucoproteínas, promueve la diferenciación celular y mantiene la integridad de los tejidos epiteliales de la piel, intestino delgado, riñones, vasos sanguíneos, útero, placenta y tracto reproductivo del macho. La vitamina A también es necesaria para la espermatogénesis, supervivencia embrionaria, crecimiento y desarrollo fetal, síntesis de glucoaminoglucanos, crecimiento de osteoclastos, hematopoyesis, desarrollo de órganos linfoides, producción de anticuerpos y respuesta inmune a los patógenos (Kang et al., 2017; Wu, 2018).

Sólo los alimentos de origen animal contienen vitamina A. Sin embargo, algunas plantas son una fuente rica en provitamina A, en forma de carotenoides. Los vegetales que son verdes oscuros, anaranjados y amarillos son buenas fuentes de carotenoides. La eficiencia de conversión de caroteno a vitamina A en el bovino es de 24% y en el ovino de 30% (Wu, 2018). Por otra parte, los alimentos expuestos al aire y a la luz solar pierden rápidamente su vitamina A. A pesar de que el pasto fresco es una excelente fuente de provitaminas A, se reduce a la mitad cuando se convierte en ensilado. También, los carotenoides y los suplementos orales que tienen vitamina A son propensos a ser destruidos en el rumen, especialmente en dietas con una alta proporción de concentrado (McDonald et al., 2010).
Vitamina D. La vitamina D está implicada en la regulación del calcio, fósforo y la hormona paratiroidea. Activa el sistema de transporte de calcio y fósforo dependiente de vitamina D en el enterocito, estimula la producción de osteoclastos para la liberación de calcio y fosfato desde el hueso, y aumenta la reabsorción de calcio y fosfato por el riñón. Así, la vitamina D es esencial para la regulación del metabolismo del calcio y fósforo, y consecuentemente, para la calcificación y crecimiento de los huesos (Nelson et al., 2016; Wu, 2018).
La vitamina D está limitada en el alimento de los animales. Raramente está presente en las plantas, excepto en forrajes secados al sol. Una dieta típica de soya y maíz no contiene vitamina D. La mayoría de animales puede sintetizar cierta cantidad de vitamina D, específicamente cuando la piel está expuesta a la luz del sol. Además, la tasa de síntesis de vitamina D en los animales varía según su ambiente. La presencia de nubes, humo o polvo reduce la radiación ultravioleta y el aire contaminado perjudica la síntesis de vitamina D en los animales en pastoreo, incluyendo vacas en lactación. Además, ya que la radiación ultravioleta no puede pasar a través de los vidrios de ventanas, los animales albergados en sistemas intensivos cerrados no pueden sintetizar vitamina D (McDonald et al., 2010; Wu, 2018).
Vitamina E. La vitamina E funciona principalmente como un antioxidante biológico en los animales. Previene la oxidación de los ácidos grasos poliinsaturados que funcionan como constituyentes primarios de las membranas celulares y los precursores de prostaglandinas, puede prevenir problemas relacionados a la oxidación tales como la infertilidad, degeneración muscular y neurológica, la disfunción cardiaca, las lesiones cutáneas y el envejecimiento. La vitamina E también desempeña un importante papel en el desarrollo y función del sistema inmune. Se ha demostrado que la suplementación con vitamina E proporciona una protección adicional contra la infección con organismos patógenos (McDonald et al., 2010; Wu, 2018).
La vitamina E no es almacenada en grandes cantidades por mucho tiempo en el cuerpo del animal y por lo tanto es necesaria una fuente regular. A pesar de que está bien distribuida en los alimentos y los forrajes son buenas fuentes de α-tocoferol, hay una pérdida de hasta el 90% de la vitamina cuando el forraje se transforma en heno. La pérdida es algo menor cuando se realiza ensilado. Durante el almacenamiento de la mayoría de granos en los silos, la actividad de la vitamina E puede declinar marcadamente (McDonald et al., 2010). La vitamina E está ampliamente distribuida en los aceites vegetales del maíz, soya, semilla de girasol, etc. Sin embargo, los granos de maíz y trigo son pobres en esta vitamina (Wu, 2018).
Vitamina B12. La vitamina B12 es sintetizada exclusivamente por microorganismos, no por las células animales. Esta vitamina no es necesaria en las dietas de los rumiantes, si ellos consumen suficiente cobalto. Las coenzimas activas de la vitamina B12 son la metilcobalamina, requerida para la síntesis de metionina, síntesis de purina, pirimidina y ácido nucleico; y la 5´-desoxy-adenosilcobalamina, requerida para la síntesis de succinil-CoA, que convierte el propionato en glucosa. La síntesis de glucosa desde propionato es importante para los rumiantes, ya que el propionato es el principal producto de la fermentación microbiana en el rumen y porque es absorbida poca glucosa desde el intestino delgado de los rumiantes (Wu, 2018).

La principal fuente de vitamina B12 son los alimentos de origen animal, siendo el hígado una fuente particularmente abundante. Se piensa que la presencia de vitamina B12 en las plantas puede deberse a una contaminación con bacterias o restos de insectos (McDonald et al., 2010). En el cuadro 1 se observa un resumen de las principales funciones de los micronutrientes revisados.
Cuadro 1. Funciones de los principales micronutrientes en rumiantes
[/et_pb_text][et_pb_image src=”https://www.corpmontana.com/wp-content/uploads/2019/04/Articulo_Extracal_1.png” align=”center” align_tablet=”center” align_phone=”” align_last_edited=”on|desktop” _builder_version=”3.23″ z_index_tablet=”500″ border_width_all=”1px”]
[/et_pb_image][et_pb_text admin_label=”Resumen” _builder_version=”3.29.3″ header_text_color=”#35568a” header_2_text_align=”left” header_3_text_color=”#35568a” header_5_text_color=”#1a9bc2″ custom_margin=”||0px|” z_index_tablet=”500″]
Consecuencias de la deficiencia de micronutrientes
Las consecuencias de la deficiencia de micronutrientes son tan variadas como sus funciones principales. Si el calcio es deficiente en la dieta de los animales jóvenes en crecimiento, entonces la formación correcta de los huesos no ocurre y se produce la condición llamada raquitismo. Los signos clínicos del raquitismo son la presentación de huesos con formas erróneas, cojeras, agrandamiento de las articulaciones y en algunos casos rigidez. En los animales adultos, la deficiencia severa de calcio produce osteomalacia, en la cual el calcio es retirado de los huesos, pero no es reemplazado (McDonald et al., 2010) y se observa principalmente después del parto en la condición llamada hipocalcemia. La hipocalcemia también puede causar paresia en los rumiantes, reflejando su papel para controlar la liberación de acetilcolina y la propagación de la señal neuromuscular. Este síndrome también es hallado en ovinos privados de alimento y que consumen pasturas conteniendo alta cantidad de oxalatos (Finnie et al., 2011).
La deficiencia de vitamina A ocurre en animales que consumen dietas poco nutritivas. Lo primero que se observa es un defecto en la visión del animal, pelo hirsuto y piel escamosa, luego se notará una disminución en la tasa de crecimiento, un incremento en el riesgo para sufrir enfermedades infecciosas, daño testicular en los machos y muerte embrionaria, reabsorción fetal y retención de placenta en las hembras. Esto ocurre cuando las reservas hepáticas de la vitamina están casi agotadas. Además, la deficiencia de vitamina A conduce a queratinización de los tejidos epiteliales del ojo, pulmones, tracto gastrointestinal y genitourinario, y disminución de la secreción mucosa. En los casos severos de deficiencia de vitamina A se observará ceguera en los animales (McDonald et al., 2010; Wu, 2018).
No hay que olvidar que la deficiencia de vitamina A en bovinos en crecimiento puede ser primaria o secundaria. La deficiencia primaria ocurre principalmente por la carencia de alimento verde durante períodos de sequía o en centros de engorde, mientras que la deficiencia secundaria ocurre por una combinación de enfermedad hepática crónica y enfermedad intestinal porque el hígado es el principal sitio de almacenamiento y la conversión de caroteno a vitamina A ocurre en el tracto alimentario. Estos casos pueden observarse en zonas con alta prevalencia de Fasciola hepatica o en zonas de pastoreo que tienen plantas tóxicas (Finnie et al., 2011; Parker et al., 2017).
Los principales signos clínicos de la deficiencia de vitamina D en animales jóvenes son las anormalidades de los huesos. Las patas pueden estar encorvadas, las articulaciones y corvejones hinchados y el lomo arqueado. Además, los huesos son débiles y estos animales pueden experimentar fracturas. En los bovinos adultos se presenta osteomalacia, principalmente en animales preñados y en lactación, que requieren cantidades grandes de calcio y fósforo (McDonald et al., 2010; Lean et al., 2013).
La manifestación más frecuente de la deficiencia de vitamina E en los animales de granja es la degeneración muscular o miopatía. La miopatía nutricional, también conocida como enfermedad del músculo blanco o distrofia muscular, frecuentemente ocurre en bovinos, particularmente terneros, cuando ellos ingieren poco pasto en el período de inverno o escasez. La miopatía afecta principalmente el músculo esquelético y los animales afectados tienen debilidad muscular de las patas, con dificultad para caminar y pararse. Eventualmente, los animales son incapaces de pararse y la debilidad de los músculos del cuello evitará que puedan levantar la cabeza. El músculo cardiaco también puede ser afectado y ello puede conducir a muerte súbita (McDonald et al., 2010; Wu, 2018).
Otras manifestaciones de la deficiencia de vitamina E incluyen la anemia en los animales debido a que se produce una disminución en la producción de hemoglobina y se acorta el ciclo biológico del eritrocito; la lipidosis hepática, desorden metabólico que se desarrolla principalmente en ovinos y caprinos cuando la disponibilidad hepática de los productos lipogénicos y glucogénicos está desbalanceada (El-Khodery et al., 2011); alteraciones de la reproducción con disminución en la producción espermática, muerte fetal, aborto espontáneo y reabsorción fetal (Wu, 2018).

Finalmente, la deficiencia de vitamina B12 es común entre los rumiantes que ingieren cantidades inadecuadas de cobalto. Asimismo, la deficiencia de esta vitamina puede ocurrir como resultado de una infección parasitaria por tenias. La deficiencia de vitamina B12 también causa una formación errónea del núcleo de los nuevos glóbulos rojos, con la consecuente acumulación de megaloblastos en la médula ósea. También se han descrito desórdenes neurológicos asociados con la deficiencia de metionina originada por la falta de vitamina B12 (Wu, 2018).
El-Khodery et al. (2011) demostraron que en cabras alimentadas con dietas conteniendo menos de 0.1 ppm de cobalto, los animales presentaron bajos niveles de vitamina B12 sérica, anemia, menores tasas de crecimiento y longitud de huesos, y una disminución en la digestibilidad aparente de nutrientes. Igualmente, McDonald et al. (2010) indican que la deficiencia de vitamina B12 en animales jóvenes causa pérdida de apetito, emaciación, anemia y alta mortalidad. En el Cuadro 2 se puede observar un resumen de los principales signos clínicos observados por la deficiencia de micronutrientes.
Cuadro 2. Signos clínicos por deficiencia de micronutrientes en rumiantes
[/et_pb_text][et_pb_image src=”https://www.corpmontana.com/wp-content/uploads/2019/04/Articulo_Extracal_2.png” align=”center” align_tablet=”center” align_phone=”” align_last_edited=”on|desktop” _builder_version=”3.23″ z_index_tablet=”500″ border_width_all=”1px”]
[/et_pb_image][et_pb_text admin_label=”Resumen” _builder_version=”4.9.0″ header_text_color=”#35568a” header_2_text_align=”left” header_3_text_color=”#35568a” header_5_text_color=”#1a9bc2″ hover_enabled=”0″ z_index_tablet=”500″ sticky_enabled=”0″]
Ventajas de la administración rutinaria de multivitamínicos
Se debe tener presente que existen algunos grupos etarios en riesgo que pueden estar propensos a experimentar una deficiencia fisiológica o adquirida de micronutrientes. Por lo tanto, es necesario tener planes de suplementación o administración parenteral de multivitamínicos para estos animales, con el fin de evitar las manifestaciones clínicas de las deficiencias. Los neonatos, por ejemplo, son íntegramente dependientes de sus madres para obtener los nutrientes vitales, tales como minerales, vitaminas e inmunoglobulinas, a través de la placenta durante el período fetal y a través del calostro y la leche luego del nacimiento (Ataollahi et al., 2018).
Se sabe que la transferencia placentaria de vitamina A al feto es limitada, por lo tanto, el neonato tiene un bajo almacenamiento de vitamina A y necesita consumir calostro en cantidades adecuadas para establecer niveles óptimos de esta vitamina (McDonald et al., 2010). La ingestión de cantidades inadecuadas de calostro o de calostro de baja calidad predisponen a la cría a tener problemas oculares. En bovinos en riesgo o que ya presentan problemas de visión por deficiencia de vitamina A, se recomienda la administración parenteral a una dosis de 400 UI/kg de peso vivo y continuar la suplementación al menos una vez al mes hasta asegurar que reciben cantidades adecuadas de vitaminas en la dieta (Parker et al., 2017).
Respecto a la vitamina D, los bovinos lecheros deben ser suplementados frecuentemente para compensar la alta demanda de esta vitamina en el período periparto para mantener la homeostasis del calcio y expresar todo el potencial de la producción láctea, ya que su concentración está en el nivel más bajo a los siete días postparto (Holcombe et al. 2018). Además, el calcio también debe ser suplementado durante la preñez y toda la lactación porque las hembras necesitan reponer diariamente el equivalente de los niveles sanguíneos circulantes de calcio para ser capaces de abastecer sus necesidades de desarrollo fetal y la síntesis de leche (Ataollahi et al., 2018). En el calostro usualmente hay seis a diez veces la cantidad de vitamina D presente en la leche motivo por el cual no es necesario un suplemento de vitamina D en el neonato (Nelson et al., 2016).
Asimismo, la vitamina D debe ser suplementada en animales de crianza intensiva cuya dieta está basada en forraje. También, ya que el contenido de vitamina D varía considerablemente en los productos como el heno y el ensilado, se recomienda la suplementación de vitamina D, especialmente para animales jóvenes y animales preñados en épocas de escasez de alimento (Martins et al., 2016; Wu, 2018).
Se han realizados muchos estudios relacionados a la suplementación y administración parenteral con multivitamínicos sobre la salud y desempeño productivo de los animales. Ataollahi et al. (2018) realizaron un estudio para determinar el efecto de la suplementación con calcio y magnesio en ovinos. Los corderos nacidos de borregas tratadas con calcio y con calcio más magnesio tuvieron una mejor respuesta contra la oxidación de leucocitos y una mejor capacidad antioxidante total en comparación a los grupos que recibieron magnesio o no recibieron suplemento. También, la producción de inmunoglobulina G en corderos mejoró desde el nacimiento hasta las cuatro semanas edad. Además, hallaron que la suplementación con estos minerales tiene un efecto significativo sobre la regulación del balance energético en borregas incrementando el nivel de calcio en leche a las cuatro semanas postparto. Ellos concluyeron que la suplementación con calcio en borregas durante la última etapa de preñez y en la lactación temprana mejora la respuesta inmune de los corderos y regula más prontamente el balance energético postparto.
Harris et al. (2018) demostraron que los bovinos que reciben una inyección de 150,000 a 300,000 UI de vitamina A al nacimiento aumentan significativamente su tasa de crecimiento y exhiben mayores pesos al destete. Un ternero destetado con más peso también requerirá menos días de engorde para alcanzar el peso final, haciendo de la vitamina A un producto potencial para incrementar la eficiencia de la producción de carne. Por otra parte, este estudio también determinó que la administración de vitamina A aumenta el depósito intramuscular de grasa, evento que está altamente relacionado a la calidad, ternura y palatabilidad de la carne.
Con relación a la vitamina D, Holcombe et al. (2018) demostraron que esta vitamina desempeña un importante papel optimizando las funciones inmunes. En vacas afectadas con mastitis, la administración de vitamina D mejoró la función inmune local, disminuyó la colonización por las bacterias que causan la mastitis y redujo la cantidad de células somáticas en leche en comparación con las vacas no tratadas. Unos años antes, Hoque et al. (2016) concluyeron que la administración de vitamina E y selenio disminuye la incidencia de mastitis y mejora el desempeño productivo y reproductivo de bovinos lecheros. Ellos utilizaron 255 vacas divididas en 5 grupos utilizando diversos esquemas de administración de vitamina E y selenio. Los grupos que recibieron vitamina E y selenio inyectable 30 días antes de la fecha probable de parto estuvieron entre 76% y 100% libres de nuevas infecciones intramamarias durante el período de lactación temprana. Se sugiere que la administración de vitamina E en el período seco incrementa la respuesta inmune a nuevas infecciones mamarias postparto.
Knowles y Grace (2016) indican que las reservas hepáticas de vitamina B12 al nacimiento son sustanciales pero los niveles disminuyen paulatinamente y al destete la concentración puede ser 30-60% menor. Lean et al. (2013) demostraron que la deficiencia de cobalto y consecuentemente de vitamina B12 está relacionada a una mayor incidencia de cojeras en los rumiantes y esto a su vez perjudica la producción láctea, pero su suplementación continua mejora los índices productivos de los animales.
En conclusión, hay muchas razones para establecer un protocolo de suplementación de multivitamínicos. Este protocolo dependerá del tipo de crianza, disponibilidad de alimento para los animales y nivel de especialización de los subproductos que se obtienen de ellos (Figura 1). Además de mejorar la rentabilidad de la crianza, el uso de suplementos multivitamínicos contribuirá con el bienestar general de los animales y la mejor calidad de productos de origen animal para los consumidores.
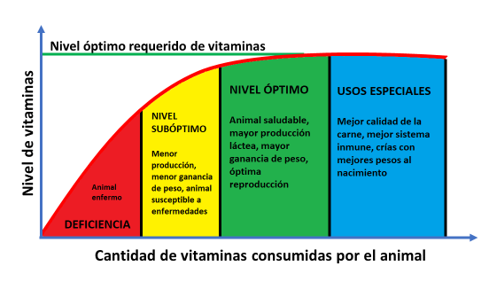
Figura 1. Niveles de vitaminas y sus consecuencias en el animal
EXTRACAL®
EXTRACAL® es un reconstituyente vitamínico que contiene cloruro de calcio, vitamina A, vitamina D3, vitamina E y vitamina B12. Está indicado para cualquier estado carencial agudo o crónico de calcio y de las vitaminas A, D, E y B12. Mejora la producción y la eficiencia reproductiva. Se recomienda como terapia adyuvante en casos de infecciones bacterianas, virales, parasitarias y metabólicas. Previene el raquitismo, osteomalacia y distrofia muscular. Indicado especialmente en la preñez y desarrollo de animales jóvenes.
Vía de administración y dosis
EXTRACAL® se administra por vía intramuscular profunda. Bovinos y equinos jóvenes: 5 ml, adultos: 10 a 12 ml. Ovinos, caprinos y camélidos sudamericanos jóvenes: 1 ml, adultos: 3 a 5 ml. Porcinos jóvenes: 0.5 a 1 ml, adultos: 5 ml. Caninos y felinos: 0.5 a 2 ml, dependiendo del tamaño y peso.
[/et_pb_text][et_pb_text admin_label=”Resumen” _builder_version=”3.27.4″ text_font_size=”11px” header_text_color=”#35568a” header_2_text_align=”left” header_3_text_color=”#35568a” header_5_text_color=”#1a9bc2″ z_index_tablet=”500″]
BIBLIOGRAFÍA
- Ataollahi F, Friend M, McGrath S, Dutton G, Peters A, Bhanugopan M. 2018. Effect of calcium and magnesium supplementation on minerals profile, immune responses, and energy profile of ewes and their lambs. Livest Sci 217: 167-173. https://doi.org/10.1016/j.livsci.2018.10.001.
- El-Khodery S, Hussein HS, El-Boshy ME, Nassif MN. 2011. Ultrasonographic evaluation to diagnose hepatic lipidosis in Egyptian Zaraibi goats with vitamin B12 deficiency. J Adv Res 2: 65-71. http://dx.doi.org/10.1016/j.jare.2010.09.004.
- Finnie JW, Windsor PA, Kessell AE. 2011. Neurological diseases of ruminant livestock in Australia. II: toxic disorders and nutritional deficiencies. Aust Vet 89: 247-253. http://dx.doi.org/10.1016/j.jare.2010.09.004.
- Harris CL, Wang B, Deavila JM, Busboom JR, Maquivar M, Parish SM, McCann B, Nelson ML, Du M. Vitamin A administration at birth promotes calf growth and intramuscular fat development in Angus beef cattle. J Anim Sci Biotechno 9: 55. https://doi.org/10.1186/s40104-018-0268-7.
- Holcombe SJ, Wisnieski L, Norby GB, Sordillo LM. Reduced serum vitamin D concentrations in healthy early-lactation dairy cattle. J Dairy Sci 101: 1488-1494. https://doi.org/10.3168/jds.2017-13547.
- Hoque MN, Das ZC, Rahman ANMA, Hoque MM. Effect of administration of vitamin E, selenium and antimicrobial therapy on incidence of mastitis, productive and reproductive performances in dairy cows. Int J Vet Sci Med 4: 63-70. http://dx.doi.org/10.1111/j.1751-0813.2011.00793.x.
- Kang S, Park C, Seo K. Ocular abnormalities associated with hypovitaminosis A in Hanwoo calves: a report of two cases. J Vet Med Sci 79(10): 1753-1756. http://dx.doi.org/10.1292/jvms.17-0166.
- Knowles SO, Grace ND. The vitamin B12 and Se status of lambs during their transition from milk-fed monogastric to grazing herbivore. New Zeal Vet J 12:35. http://dx.doi.org/10.1080/00480169.2016.1252701.
- Lean IJ, Westwood CT, Golder HM, Vermunt JJ. Impact of nutrition on lameness and claw health in cattle. Livest Sci 156: 71-87. http://dx.doi.org/10.1016/j.livsci.2013.06.006.
- Martins ERM, Bertagnon HG, Batista CF, Gomes RC, Santos KR, Bellinazzi JB, Della Libera AMMP. Influência da suplementaҫão de vitaminas A, D e E na funҫão imune de bezerros alimentados com dieta á base de feno capim tifton (Cynodon spp.). Pesq. Vet. Bras. 36(5): 453-459. http://dx.doi.org/10.1590/S0100-736X2016000500016.
- McDonald P, Edwards RA, Greenhalgh JFD, Morgan CA, Sinclair LA, Wilkinson RG. 2010. Animal nutrition. 7ª ed. Harlow: Pearson. 714 p.
- Nelson CD, Lippolis JD, Reinhardt TA, Sacco RE, Powell JL, Drewnoski ME, O’Neil DC, Weiss WP. 2016. Vitamin D status of dairy cattle: outcomes of current practices in the dairy industry. J Dairy Sci 99: 1-11. http://dx.doi.org/10.3168/jds.2016-11727.
- Parker EM, Gardiner CP, Kessell AE, Parker AJ. Hypovitaminosis A in extensively grazed beef cattle. Aust Vet J 95(3): 80-84. http://dx.doi.org/10.1111/avj.12560.
Wu G. 2018. Principles of animal nutrition. Filadelfia: CRC Press. 801 p.
[/et_pb_text][/et_pb_column][/et_pb_row][/et_pb_section]